B형 간염 바이러스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
B형 간염 바이러스는 헤파드나바이러스과에 속하는 DNA 바이러스로, 간염, 간경변, 간세포암을 유발한다. 1883년 독일에서 발생한 집단 발병 사례를 통해 처음 기록되었으며, 1964년 배리 블루버그가 오스트레일리아 항원을 발견하면서 연구가 시작되었다. 바이러스는 지질 외피와 단백질로 구성된 42nm 크기의 데인 입자 형태로, 유전체는 원형 DNA로 구성된다. B형 간염 바이러스는 혈액을 통해 감염되며, 수직 감염(출산 시 산모에서 신생아로의 전파)과 수평 감염(성관계, 주사기 공유 등) 경로가 있다. 전 세계적으로 유행하며, 특히 아시아, 아프리카 등에서 만연하며, 치료를 통해 간 손상을 줄이고 합병증 발생을 예방하는 것을 목표로 한다.

더 읽어볼만한 페이지
- B형 간염 - B형 간염 백신
B형 간염 백신은 B형 간염 바이러스 감염을 예방하기 위해 사용되며, 신생아에게 출생 시 예방 접종을 권장하고, 면역 기억을 통해 최소 30년 동안 보호 효과를 제공하며, 심각한 부작용은 드물다.

2. 역사
B형 간염 바이러스(HBV)에 관한 최초의 기록은 1883년 독일 제국의 루어만이 보고한 것이다.[68] 당시 사람의 림프액이 함유된 천연두 백신을 접종받은 독일 조선소 노동자들에게서 간염 증상이 나타났는데, 이는 현재 B형 간염으로 알려져 있다.[68] 1943년 폴 비슨(Paul Beeson)은 수혈을 받은 환자들에게서 황달이 발생하는 것을 묘사하면서 혈액을 통한 B형 간염 바이러스 전파를 강조하였다. 1947년에는 A형 간염 바이러스와 B형 간염 바이러스의 차이가 밝혀졌다.[69]
1964년 미국의 배리 블루버그는 오스트레일리아 원주민의 혈청에서 "오스트레일리아 항원"을 발견했는데,[70] 이는 훗날 B형 간염 바이러스 표면 항원(HBsAg)으로 확인되었다. 1968년 도쿄 대학 의학부의 오코치 카즈오(후에 규슈 대학 의학부 교수)는 오스트레일리아 항원과 간염의 관련성을 보고했다.
분자생물학적 연구를 통해, 독일 뮌스터 대학교 연구팀은 조류의 DNA에서 B형 간염 바이러스의 흔적을 발견했다. 이를 통해 조류 감염은 8200만 년 전부터, 포유류 감염은 1210만 년 이후부터 시작되었을 것으로 추정된다.[83]
3. 구조
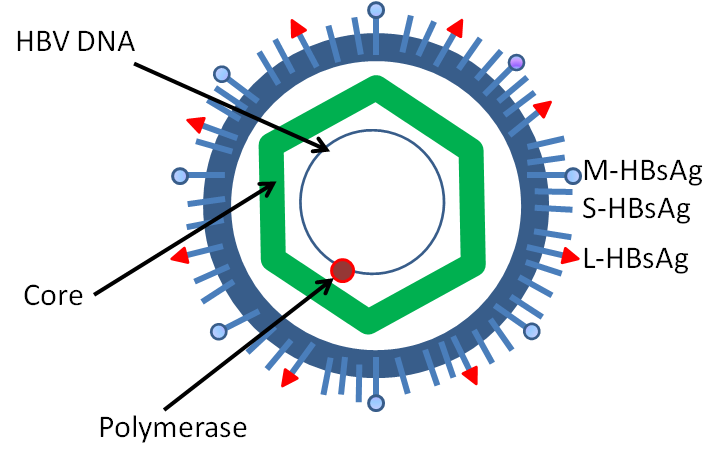
B형 간염 바이러스는 직경 42nm의 구형 입자로, 외피(envelope)와 뉴클레오캡시드(nucleocapsid)로 구성된다. 외피는 지질과 단백질(HBsAg)로 이루어져 있으며, 바이러스가 숙주 세포에 부착하고 침투하는 데 관여한다. 뉴클레오캡시드는 정이십면체 모양이며, 내부에는 바이러스 DNA와 DNA 중합효소(polymerase)가 들어있다.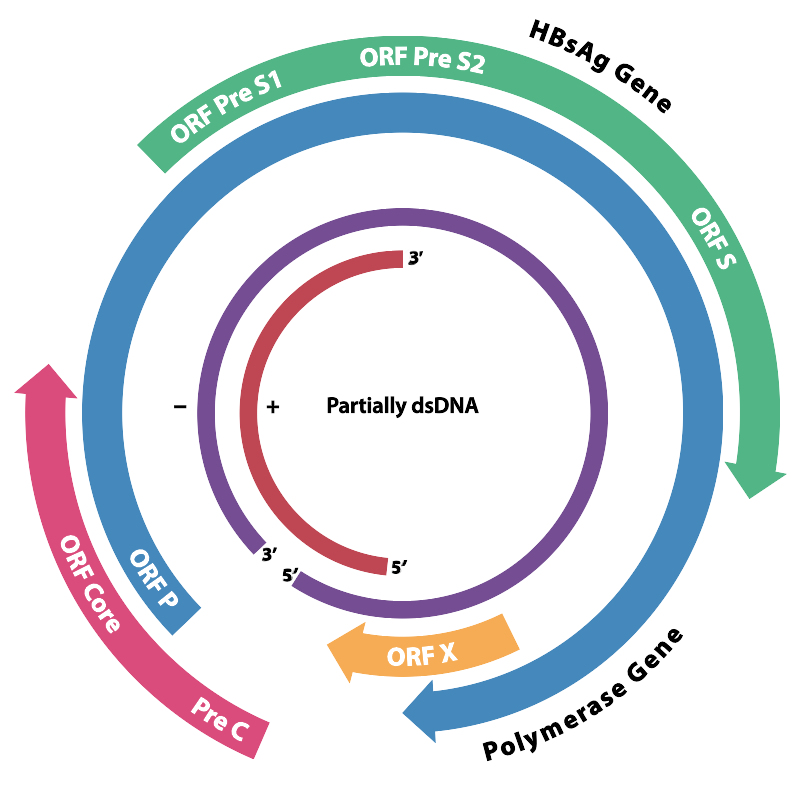
바이러스 DNA는 약 3.2kb 크기의 불완전한 이중 가닥 환형 DNA이다. 완전한 길이의 가닥 한쪽 끝은 바이러스 DNA 중합효소에 연결되어 있다. 유전체 길이는 3020~3320 뉴클레오타이드(전체 길이 가닥의 경우)이며, 1700~2800 뉴클레오타이드(짧은 길이 가닥의 경우)이다.[34] 표면에 있는 항원 (HBs)에 따라 adr, adw, ayr, ayw 등의 아형이 존재한다.
3. 1. 구성 요소
B형 간염 바이러스는 헤파드나바이러스과에 속하며, 외부 지질 외피와 단백질로 구성된 정이십면체 뉴클레오캡시드 코어로 이루어진 입자(데인 입자[11])로 구성된다.[10] 뉴클레오캡시드는 바이러스 DNA와 역전사 효소 활성을 가진 DNA 중합효소를 포함한다.[12] 외부 외피에는 바이러스가 세포에 결합하고 침투하는 데 필요한 단백질이 있다. B형 간염 바이러스는 가장 작은 외피를 가진 동물 바이러스 중 하나(비리온 직경 42nm)이지만, 코어가 없는 필라멘트 형태 및 구형체 등 다형성 형태도 존재한다. 이러한 입자들은 감염성이 없으며, 표면 항원(HBsAg)이라고 불리는 지질과 단백질로 구성되어 바이러스 생애 주기 동안 과잉 생산된다.[13]
B형 간염 바이러스는 다음과 같은 구성 요소로 이루어져 있다.
D형 간염 바이러스는 독성을 나타내기 위해 B형 간염 바이러스 외피 입자가 필요하다.[25]
4. 분류
B형 간염 바이러스(HBV)는 헤파드나바이러스과(Hepadnaviridae) 오르토헤파드나바이러스(Orthohepadnavirus) 속에 속하며, 이 속에는 B형 간염 바이러스 외에 11종의 다른 종들이 포함되어 있다.[91][3] 헤파드나바이러스과에는 아비헤파드나바이러스(Avihepadnavirus) 속도 포함된다.[91][3]
유인원(오랑우탄, 긴팔원숭이, 보노보, 고릴라, 침팬지), 구세계 원숭이,[4] 신세계 원숭이의 양털 원숭이(양털 원숭이 B형 간염 바이러스)에서 B형 간염과 유사한 바이러스가 발견되어, 이 바이러스가 영장류에서 기원했을 가능성이 제기되었다.
B형 간염 바이러스는 외피 단백질에 존재하는 항원 결정기에 따라 4가지 주요 항원형(adr, adw, ayr, ayw)으로 분류된다.
4. 1. 유전자형
B형 간염 바이러스는 항원 에피토프가 외피 단백질에 존재하는 것을 기반으로 네 가지 주요 혈청형(adr, adw, ayr, ayw)으로 나뉜다. 이러한 혈청형은 공통 결정인 (a)와 두 개의 상호 배타적 결정인 쌍 (d/y 및 w/r)을 기반으로 한다. 바이러스 균주는 게놈의 전체 뉴클레오티드 서열 변이에 따라 10개의 유전자형(A-J)과 40개의 아형으로 나뉘었다.[5] 이 유전자형은 뚜렷한 지리적 분포를 보이며 바이러스의 진화와 전파를 추적하는 데 사용된다. 유전자형 간의 차이는 질병의 심각도, 경과 및 합병증의 가능성, 치료 반응에 영향을 미친다.[6][7] 혈청형과 유전자형이 반드시 일치하는 것은 아니다.유전자형은 염기 서열에서 최소 8%의 차이를 보이며, 뚜렷한 지리적 분포를 보인다. 유전자형 내에는 아형이 설명되어 있으며, 이들은 유전체에서 4~8%의 차이를 보인다.
A부터 J까지 10개의 알려진 유전자형이 있다.
;개별 유전자형
F형은 다른 유전자형과 14% 차이를 보이는 가장 분기된 유형이다.
E 및 F 계통은 각각 아프리카와 신세계의 원주민에서 유래된 것으로 보인다.
5. 생활환(Life cycle)
B형 간염 바이러스는 복잡한 생활 주기를 거친다. B형 간염 바이러스는 레트로바이러스는 아니지만, 복제 과정의 일부로 역전사를 사용하는 몇 안 되는 바이러스 중 하나이다.
- 부착: 바이러스는 세포 표면의 수용체에 결합하고, 클라트린이나 caveolin-1이 매개하는 세포내이입을 통해 세포 안으로 들어간다.[50] B형 간염 바이러스는 우선 헤파린 황산염 프로테오글리칸에 결합한다. 이후 B형 간염 바이러스 L 단백질의 pre-S1 부분이 ''SLC10A1'' 유전자에 의해 암호화된 세포 표면 수용체인 나트륨 타우로콜레이트 공수송 폴리펩타이드(NTCP)에 강하게 결합한다.[51] NTCP는 주로 간세포의 동맥 혈관막에서 발견된다. 간세포 내 NTCP의 존재는 B형 간염 바이러스 감염의 조직 특이성과 관련이 있다.[50]
- 침투: 세포내이입 후, 바이러스 막은 숙주 세포의 막과 융합되어 핵캡시드를 세포질로 방출한다.[52]
- 탈피: 바이러스는 숙주 효소에 의해 생성된 RNA를 통해 증식하기 때문에, 바이러스 유전체 DNA는 세포 핵으로 전달되어야 한다. 캡시드는 미세소관을 통해 핵공으로 이동하는 것으로 추정된다. 코어 단백질은 부분적으로 이중 가닥인 바이러스 DNA에서 분리된 다음, (숙주 DNA 중합효소에 의해) 완전한 이중 가닥으로 만들어지고 4개의 바이러스 mRNA 전사의 템플릿 역할을 하는 공유 결합 폐쇄 원형 DNA(cccDNA)로 변환된다.
- 복제: 가장 큰 mRNA (바이러스 유전체보다 더 길다)는 유전체의 새로운 복사본을 만들고, 캡시드 코어 단백질과 바이러스 RNA 의존 DNA 중합효소를 만드는 데 사용된다.
- 조립: 이 4개의 바이러스 전사체는 추가 처리를 거쳐 세포에서 방출되거나 핵으로 반환되어 훨씬 더 많은 복사본을 생성하기 위해 재활용되는 자손 바이리온을 형성한다.[38][53]
- 방출: 긴 mRNA는 다시 세포질로 수송되어 바이리온 P 단백질이 역전사 효소 활성을 통해 DNA를 합성한다.
B형 간염 바이러스는 DNA 바이러스이다. 지름 42nm로 3.2kb의 환상 이중 가닥 DNA와 이를 둘러싼 외피로 구성된다. 다만, 이 DNA는 완전한 이중 가닥이 아니라 플러스 가닥 쪽이 일부 짧게 결여되어 있다. 내부에는 코어(Dane 입자라고도 함)를 가지고 있다. 표면에 있는 항원 (HBs)에 따라 adr, adw, ayr, ayw 등의 아형이 존재한다.
B형 간염 바이러스(HBV)가 세포에 감염되면 먼저 자신의 DNA를 세포의 핵으로 보낸다. 거기서 플러스 가닥 DNA를 복구하여 완전한 이중 가닥의 슈퍼코일 DNA가 되면 이를 주형으로 하여 숙주 세포 유래의 RNA 중합 효소를 사용하여 프리게놈 RNA를 합성한다. 거기에서 바이러스 DNA 중합 효소에 있는 역전사 효소 활성을 이용하여 이 RNA로부터 마이너스 가닥 DNA를 합성한다. 이 과정에서는 RNA의 프라이머가 차례로 전이되어 전체 길이의 마이너스 가닥 DNA를 완성시킨다. 그 후, DNA 중합 효소에 의해 플러스 가닥이 합성되지만, 합성이 모두 끝나기 전에 소포체 내강으로 출아한 바이러스 표면 분자인 HBs에 덮여 세포 외로 방출된다.
B형 간염 바이러스는 열에 강하여 60℃ × 10분간의 가열 처리로도 불활성화되지 않고 감염성을 잃지 않지만[84], 60℃ × 10시간에서는 불활성화된다.[85]
6. 질병
B형 간염 바이러스(HBV)는 간염, 간경변, 간세포암 등 다양한 간 질환을 일으킬 수 있으며, 췌장암 위험 증가와도 관련이 있다는 보고가 있다.[90][56][89][55]
HBV 감염은 급성으로 시작해 만성으로 진행될 수 있으며, 이 과정에서 다른 질병이나 건강 상태를 유발하기도 한다.[54]
6. 1. 질병의 역할
HBx 단백질은 세포 경로 조절, DNA 결합, miRNA 발현 패턴 변화 등을 통해 간세포에 다양한 변화를 유발한다. B형 간염 바이러스 감염은 세포 내 활성산소(ROS)를 증가시켜 산화적 DNA 손상을 일으키고, 이는 돌연변이 및 후성유전적 변화를 유발하여 간 질환 및 간암 발생에 기여할 수 있다.[57][60][61]HBx는 만성 B형 간염 감염 시 세포 내 활성산소(ROS)가 약 10,000배 증가하는 것의 부분적인 원인이다. ROS 증가는 부분적으로 HBx가 미토콘드리아에 국한되어 미토콘드리아 막 전위를 감소시키기 때문에 발생할 수 있다.[58] 또한, 다른 B형 간염 바이러스 단백질인 HBsAg도 소포체와의 상호 작용을 통해 ROS를 증가시킨다.[58]
B형 간염 바이러스 감염 후 ROS 증가는 염증을 유발하고, 이는 ROS의 추가적인 증가로 이어진다. ROS는 20가지 이상의 DNA 손상을 유발한다.[59] 산화적 DNA 손상은 돌연변이를 일으키며, DNA 손상 복구 과정에서 손상 부위에 후성 유전적 변화가 일어날 수 있다.[60][61]
후성 유전적 변화와 돌연변이는 간 질환에 기여하는 세포 기계 장치의 결함을 유발할 수 있다. 축적된 후성 유전적 및 돌연변이적 변화가 결국 암으로 진행될 때, 후성 유전적 변화는 이러한 암 발생에서 돌연변이보다 더 큰 역할을 하는 것으로 보인다. TP53[62]와 ARID1A[63] 유전자 정도만이 간암의 20% 이상에서 돌연변이가 발생하는 반면, 41개의 유전자는 각각 과메틸화된 프로모터(유전자 발현을 억제)를 간암의 20% 이상에서 가지고 있으며, 이 중 7개의 유전자는 간암의 75% 이상에서 과메틸화되어 있다.[62]
DNA 복구 부위 변화 외에도, 후성 유전적 변화는 HBx가 DNA 메틸트랜스퍼라제 효소인 DNMT1 및/또는 DNMT3A를 특정 유전자 부위에 모집하여 해당 유전자의 메틸화 수준과 유전자 발현을 변화시킴으로써 유발된다.[64] HBx는 또한 유전자 발현에 영향을 줄 수 있는 히스톤 아세틸화 및 탈아세틸화를 변화시킨다.[64]
수천 개의 단백질 코딩 유전자가 HBx 결합 부위를 갖는 것으로 보이며,[57][65] 단백질 코딩 유전자 외에도, 약 15개의 마이크로RNA와 16개의 긴 비암호 RNA도 HBx가 프로모터에 결합함으로써 영향을 받는다.[65]
7. 감염
B형 간염 바이러스는 주로 혈액을 통해 감염된다. 감염 경로는 수직 감염과 수평 감염이 있으며, 성인 이후의 수평 감염은 일과성인 경우가 많다.
수직 감염은 출산 과정에서 산모로부터 신생아에게 전파되는 경우가 가장 흔하다. 수평 감염은 감염된 사람과의 성관계, 오염된 주사기 공동 사용, 날카로운 물체에 노출되는 경우 발생할 수 있다. 일회용이 아니거나 오토클레이브(고압 증기 멸균기)로 제대로 소독되지 않은 바늘은 어떤 종류든 위험할 수 있으며, 여기에는 문신 및 피어싱 시술소에서 사용되는 바늘도 포함된다.[75] 귀걸이나 기타 신체 피어싱 장신구를 공유하는 것도 B형 간염 바이러스 전파의 원인이 될 수 있다.[76]
HBeAg 양성 환자가 사용한 혈액 투석 장치는 바이러스 전파 위험이 높다. 혈액 투석 장치는 일반적으로 여러 환자를 동시에 치료하므로 혈액 오염이 발생할 수 있기 때문이다. 미국에서는 혈액 투석 장치를 통한 HBV 감염 발생률이 1% 수준이다. 의료 종사자 역시 감염 위험이 높다.[79][80] B형 간염 백신 접종은 HBV 전파를 억제하는 효과적인 방법이지만, 바이러스 유행 지역, 특히 의료 시설이 부족한 시골 지역에서는 백신 공급이 제한적이다.
HBV는 표면에서 최대 7일 동안 감염성을 유지할 수 있다. 그러나 모유 수유, 식기 공유, 포옹, 키스, 손 잡기, 기침, 재채기 등으로는 전파되지 않으며, 다른 간염 바이러스와 달리 오염된 음식이나 물을 통해서도 전파되지 않는다. 다만, B형 간염 바이러스 감염자와 함께 생활하는 경우 감염 위험이 높아진다.[68]
8. 역학
B형 간염 감염률은 남성과 여성 모두 동일하다. B형 간염 바이러스는 아시아, 태평양 섬 주민, 또는 아프리카계 미국 시민에게서 더 두드러지게 발견되며, 이들 중 약 25%가 진단을 받게 된다.[73] B형 간염 바이러스는 정맥 주사 약물 사용, 복수의 성 파트너, 남성 동성애자 등과 같이 위험 행동을 하는 집단에서 더 쉽게 전파된다.[74]
B형 간염 바이러스는 전 세계 거의 모든 지역에서 발견될 수 있지만, 아시아, 아프리카, 남아메리카, 카리브해에 위치한 일부 국가에서 만연해 있다.[75]
약 20억 명의 사람들이 B형 간염 바이러스에 감염되었으며, 이는 거의 3명 중 1명이 감염되었다는 것을 의미한다. 매년 약 150만 명이 새롭게 감염되며, 이들 중 약 10%는 진단을 받지 못한다. 매년 약 82만 명이 B형 간염 감염 및 관련 합병증으로 사망한다.[76]
임신 중 B형 간염 바이러스의 전파는 어린 시절에 만성 B형 간염을 유발하는 가장 높은 위험 요인으로 남아 있다. 감염된 유아의 약 90%가 만성적으로 감염된다. B형 간염 바이러스에 감염된 성인의 2%-6%만이 만성적으로 감염된다.[77] 전 세계적으로 B형 간염 바이러스에 만성적으로 감염된 약 3억 5천만 명 중 50% 이상이 출생 전 또는 어린 시절에 감염되었다. B형 간염 바이러스가 만연한 국가에서는 가임기 여성의 HBeAg 양성률이 높아 신생아에게 B형 간염 바이러스를 전파할 수 있기 때문에 B형 간염 바이러스의 수직 감염이 주요 건강 위험으로 작용한다. B형 간염 바이러스가 만연한 지역에서는 감염이 고위험 행동 집단에만 국한되지 않는다. 대신, 감염은 다양한 전파 경로를 통해 발생하지만, 주로 어린 시절에 발생한다.[78]
서구 세계에서 B형 간염 바이러스의 확산은 주로 성관계 또는 정맥 주사 약물 사용자(IVDU)의 주사기 공유를 통해 발생한다. IVDU는 유럽과 북미에서 B형 간염 바이러스 감염률이 가장 높다.[74] 남성 동성애자(MSM) 사이에서도 B형 간염 감염률이 더 높다.
9. 치료
B형 간염 치료의 목적은 만성 간염의 진정(ALT의 정상화)과 그 후의 간경변, 간세포암종 발병을 막는 것이다. 급성 B형 간염은 기본적으로 보존적 치료가 이루어진다.
10. 동시 감염
B형 간염은 C형 간염, D형 간염(B형 간염의 위성 바이러스), 인간 면역결핍 바이러스(HIV) 등과 같은 다른 바이러스와 함께 한 개인에게 동시 감염될 수 있다.
HBV와 HCV는 전파 방식이 비슷하여 동시 감염이 가능하다. HBV와 HCV의 동시 감염은 주로 정맥 주사 약물 사용자, 선별되지 않은 혈액 제제, 오염된 주사 바늘 및 멸균되지 않은 의료 장비 노출을 통해 발생한다. 이 두 바이러스의 동시 감염은 더 심각한 간 질환을 유발하고 원발성 간암(간세포암)의 위험을 증가시킬 수 있다. C형 간염이 동시 감염 동안 우세한 간 바이러스가 되어 신체에서 검출 가능한 HBV의 양을 감소시키기 때문에, 이러한 동시 감염 보고가 과소 보고될 수 있다.[81] 최근 통계에 따르면 HIV 감염자의 10%가 B형 간염에도 감염되어 있으며, 동남아시아의 경우 이 수치는 거의 20%까지 증가한다. B형 간염 감염은 항레트로바이러스 치료법 개발 및 사용 이후 HIV 환자에게서 입원 및 사망의 주요 원인 중 하나이다. HIV와 HBV에 모두 감염된 사람은 만성 B형 간염에 걸릴 확률이 6배 더 높다. 일부 연구에서는 이는 동시 감염된 개인의 CD4+ T 세포 수가 더 낮기 때문일 수 있다고 제안한다.[82]
참조
[1]
서적
Molecular Virology of Human Pathogenic Viruses
Academic Press
[2]
웹사이트
Hepatitis viruses
http://pathmicro.med[...]
University of Southern California, Department of Pathology and Microbiology
2007-11-21
[3]
웹사이트
ICTV Report Hepadnaviridae
http://www.ictv.glob[...]
[4]
논문
Discovery of naturally occurring transmissible chronic hepatitis B virus infection among Macaca fascicularis from Mauritius Island.
2013-11
[5]
논문
A novel hepatitis B virus subgenotype D10 circulating in Ethiopia
2017-02
[6]
논문
Hepatitis B virus genotypes
2005-03
[7]
논문
Subtypes, genotypes and molecular epidemiology of the hepatitis B virus as reflected by sequence variability of the S-gene
[8]
논문
New HBV subgenotype D9, a novel D/C recombinant, identified in patients with chronic HBeAg-negative infection in Eastern India
2013-03
[9]
논문
Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes
2013-10
[10]
서적
Baron's Medical Microbiology
Univ of Texas Medical Branch
2018-04-11
[11]
웹사이트
WHO {{!}} Hepatitis B
https://www.who.int/[...]
2015-07-12
[12]
논문
Molecular virology of hepatitis B virus
[13]
논문
The biology of hepadnaviruses
1986-07
[14]
논문
Hepatitis B surface antigen (HBsAg) levels in the natural history of hepatitis B virus (HBV)-infection: a European perspective
2010-04
[15]
논문
Hepatitis B virus biology
2000-03
[16]
논문
Hepatitis B virus nucleocapsid but not free core antigen controls viral clearance in mice
2012-09
[17]
논문
Hepatitis B virus core antigen determines viral persistence in a C57BL/6 mouse model
2010-05
[18]
논문
A mutant hepatitis B virus core protein mimics inhibitors of icosahedral capsid self-assembly
2009-03
[19]
논문
Nucleoside/nucleotide analog inhibitors of hepatitis B virus polymerase: mechanism of action and resistance
2014-10
[20]
논문
Persistence of hepatitis B virus covalently closed circular DNA in hepatocytes: molecular mechanisms and clinical significance
2014-09
[21]
웹사이트
TSRI - News and Publications
http://www.scripps.e[...]
2009-01-03
[22]
논문
Molecular functions and biological roles of hepatitis B virus x protein
2006-10
[23]
논문
Hepatitis B virus replication is associated with an HBx-dependent mitochondrion-regulated increase in cytosolic calcium levels
2007-11
[24]
논문
Activation and inhibition of cellular calcium and tyrosine kinase signaling pathways identify targets of the HBx protein involved in hepatitis B virus replication
2003-07
[25]
논문
Properties of subviral particles of hepatitis B virus
2008-08
[26]
논문
The genome of a Mesozoic paleovirus reveals the evolution of hepatitis B viruses
2013-04-30
[27]
논문
Bat hepadnaviruses and the origins of primate hepatitis B viruses
2016-02
[28]
논문
A vestigial X open reading frame in duck hepatitis B virus
[29]
논문
Ten millennia of hepatitis B virus evolution
https://www.science.[...]
2021-10-08
[30]
논문
Ancient hepatitis B viruses from the Bronze Age to the Medieval period
http://urn.kb.se/res[...]
2018-05
[31]
논문
Neolithic and medieval virus genomes reveal complex evolution of hepatitis B
2018-05-10
[32]
논문
Dating the origin and dispersal of hepatitis B virus infection in humans and primates
2013-03
[33]
논문
Tracing hepatitis B virus (HBV) genotype B5 (formerly B6) evolutionary history in the circumpolar Arctic through phylogeographic modelling
2017
[34]
논문
Hepatitis B virus genetic variability and evolution
http://www.hal.inser[...]
2007-08
[35]
논문
Enhancer I predominance in hepatitis B virus gene expression
2004-02
[36]
논문
Hepatitis B virus (HBV) promoters are regulated by the HBV enhancer in a tissue-specific manner
1989-02
[37]
논문
Regulation of hepatitis B virus gene expression
[38]
논문
Hepatitis B virus replication
2007-01
[39]
논문
The enigmatic X gene of hepatitis B virus
2004-12
[40]
논문
A long HBV transcript encoding pX is inefficiently exported from the nucleus
2003-05
[41]
논문
The hepatitis B virus post-transcriptional regulatory element contains two conserved RNA stem-loops which are required for function
1998-11
[42]
논문
The apical stem-loop of the hepatitis B virus encapsidation signal folds into a stable tri-loop with two underlying pyrimidine bulges
2002-11
[43]
논문
Possible new hepatitis B virus genotype, southeast Asia
2008-11
[44]
논문
When should "I" consider a new hepatitis B virus genotype?
2008-08
[45]
논문
Full-genome sequence of a hepatitis B virus genotype f1b clone from a chronically infected chilean patient
2014-10
[46]
논문
Hepatitis B genotypes and response to antiviral therapy: a review
[47]
논문
Hepatitis B virus genotypes: an overview
http://www.hbpdint.c[...]
2008-10
[48]
논문
A new isolate of hepatitis B virus from the Philippines possibly representing a new subgenotype C6
2009-06
[49]
논문
Novel subgenotypes of hepatitis B virus genotypes C and D in Papua, Indonesia
2008-07
[50]
논문
Visualization of hepatitis B virus entry - novel tools and approaches to directly follow virus entry into hepatocytes
2016-07
[51]
논문
NTCP opens the door for hepatitis B virus infection
2015-09
[52]
논문
Hepatitis B Virus and Hepatitis D Virus Entry, Species Specificity, and Tissue Tropism
2015-08
[53]
논문
Hepatitis B virus morphogenesis
2007-01
[54]
논문
Revisiting Hepatitis B Virus: Challenges of Curative Therapies
[55]
논문
Solution structure of stem-loop alpha of the hepatitis B virus post-transcriptional regulatory element
2008-03
[56]
논문
Association between hepatitis B virus and pancreatic cancer
2008-10
[57]
논문
Epigenetic Regulation of Viral Biological Processes
2017-11
[58]
논문
'Liver let die': oxidative DNA damage and hepatotropic viruses
http://pure-oai.bham[...]
2014-05
[59]
논문
Occurrence, Biological Consequences, and Human Health Relevance of Oxidative Stress-Induced DNA Damage
2016-12
[60]
논문
Oxidatively induced DNA damage: mechanisms, repair and disease
2012-12
[61]
논문
Oxidative stress and epigenetic instability in human hepatocarcinogenesis
[62]
논문
Genetics and epigenetics of liver cancer
2013-05
[63]
논문
Exploration of liver cancer genomes
2014-06
[64]
논문
Hepatitis B virus X protein-induced aberrant epigenetic modifications contributing to human hepatocellular carcinoma pathogenesis
2013-08
[65]
논문
Genome-wide identification of direct HBx genomic targets
2017-02
[66]
논문
Hepatitis B: The virus and disease
2009-05
[67]
웹사이트
VA.gov {{!}} Veterans Affairs
https://www.hepatiti[...]
2022-12-04
[68]
웹사이트
Pinkbook: Hepatitis B {{!}} CDC
https://www.cdc.gov/[...]
2022-12-03
[69]
논문
A historical perspective on the discovery and elucidation of the hepatitis B virus
https://www.scienced[...]
2016-07-01
[70]
논문
Polymorphisms of the serum proteins and the development of iso-preciptins in transfused patients
[71]
논문
ENGERIX-B
https://www.fda.gov/[...]
2019-10-03
[72]
웹사이트
Immunization Schedules for Heplisav-B Vaccine
https://www.cdc.gov/[...]
2022-12-04
[73]
웹사이트
Hepatitis B Facts and Figures
https://www.hepb.org[...]
2022-12-02
[74]
논문
Hepatitis B: transmission by sexual contact and needle sharing
https://dx.doi.org/1[...]
1990-03-01
[75]
웹사이트
Hepatitis B
https://wwwnc.cdc.go[...]
2022-12-02
[76]
웹사이트
Hepatitis B
https://www.who.int/[...]
2022-12-02
[77]
웹사이트
Safety Information for Hepatitis B Vaccines
https://www.cdc.gov/[...]
2022-12-04
[78]
논문
Epidemiological Patterns of Hepatitis B Virus (HBV) in Highly Endemic Areas
1996
[79]
논문
Clinical practice guideline management of blood borne viruses within the haemodialysis unit
2019-10-28
[80]
논문
Transmission of Hepatitis B Virus in Dialysis Units: A Systematic Review of Reports on Outbreaks
http://journals.sage[...]
2015-01
[81]
웹사이트
Hepatitis B Foundation: Hepatitis C and Hepatitis B Coinfection
https://www.hepb.org[...]
2022-12-02
[82]
논문
Hepatitis B and human immunodeficiency virus coinfection
2009-05
[83]
뉴스
8200万年前に鳥類感染か=B型肝炎ウイルス―独大学
http://www.jiji.com/[...]
時事通信
2013-05-01
[84]
간행물
B型肝炎ウイルスの不活化
https://doi.org/10.2[...]
1977
[85]
간행물
加熱人血漿蛋白液輸液とHBe抗原・抗体
https://doi.org/10.2[...]
1978
[86]
웹사이트
Mortality and Burden of Disease Estimates for WHO Member States in 2002
http://www.who.int/e[...]
World Health Organization
2009-11-13
[87]
서적
Molecular Virology of Human Pathogenic Viruses
https://archive.org/[...]
Academic Press
[88]
웹인용
Hepatitis viruses
http://pathmicro.med[...]
University of Southern California, Department of Pathology and Microbiology
2007-11-21
[89]
저널
Solution structure of stem-loop alpha of the hepatitis B virus post-transcriptional regulatory element
2008-03
[90]
저널
Association between hepatitis B virus and pancreatic cancer
2008-10
[91]
웹인용
Virus Taxonomy: 2018b Release
https://talk.ictvonl[...]
2019-02
[92]
웹인용
00.030. Hepadnaviridae
https://www.ncbi.nlm[...]
International Committee on Taxonomy of Viruses
2008-07-08
[93]
뉴스
Discovery of naturally occurring transmissible chronic hepatitis B virus infection among Macaca fascicularis from Mauritius Island.
2013-11
[94]
저널
Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes
2013-10
[95]
웹인용
Fam46A (Protein Coding)
https://www.genecard[...]
GeneCards
2015-02-18
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com